Research Highlights 2010
Fluidization of Tissues by Cell Division and Apoptosis
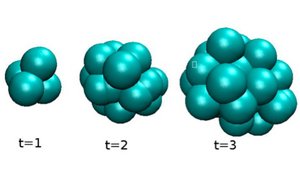
When a cell divides in a tissue, stresses are locally generated which induce displacements of other cells in the surrounding volume. These stresses are anisotropic and typically correspond to expansion along the cell division axis. Here we show that repeated cell division and cell death can lead to the relaxation of elastic stresses during a characteristic time. As a consequence, an arrangement of cells with solid material properties can behave as a viscous fluid on long times it is undergoes repeated cell division and/or cell death.
J. Ranft, M. Basan, J. Elgeti, J.-F. Joanny, J. Prost and F. Jülicher
Proc. Natl. Acad. Sci. USA 107, 20863 (2010)
[PDF (284 kB)]; Supplementary Material
Anisotropies in Cortical Tension Reveal the Physical Basis of Polarizing Cortical Flows
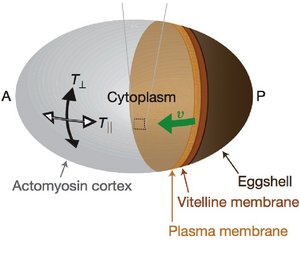
Associated with the cell membrane is a thin layer of a gel-like cytoskeletal network, the cell cortex. The cell cortex is an active gel in which myosin motor molecules can generate
spontaneous flow patterns. Using a combination of experiments and theory, we show that the observed flow patterns can be understood as the consequence of force balances involving active and viscous stresses in the material.
M. Mayer, M. Depken, J. Bois, F. Jülicher and S. Grill
Nature 467, 617 (2010)
[PDF (952 kB)]
Collective Behavior of Antagonistically Acting Kinesin-1 Motors
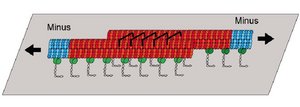
Molecular motors that cooperate in large groups show collective behaviors such as bidirectional motion. We show that the movements generated by antagonistic groups of motors observed in motility assays with antiparallel filaments can be understood quantitatively by a simple theory. The quantitative comparison between experiment and theory reveals the importance of force-dependent detachment of motors and highlights the role elastic elements linking the motor to the substrate.
C. Leduc, N. Pavin, F. Jülicher and S. Diez
Phys. Rev. Lett. 105, 128103 (2010)
[PDF (648 kB)]
Cell Flow Reorients the Axis of Planar Polarity in the Wing Epithelium of Drosophila
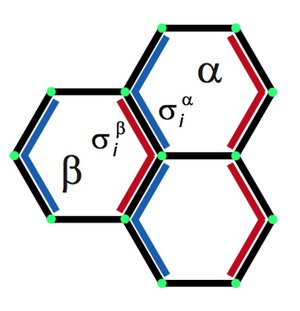
Two dimensional tissues exhibit large scale polarity patterns which govern for example the orientation of hair-like structures. Here we show that the large scale reorientation of polarity during pupal development of the fly wing is guided by cell flow patterns which result from active tissue remodeling. Based on quantitative observations we discuss key physical mechanisms that are involved in polarity reorganization. We propose a simple vertex model to describe the interplay of cell polarity with rearrangements of the cellular network.
B. Aigouy, R. Farhadifar, D. B. Staple, A. Sagner, J.-C. Röper, F. Jülicher and S. Eaton
Cell 142, 773 (2010)
[PDF (3,5 MB)]
The Interplay between Active Hair Bundle Motility and Electromotility in the Cochlea
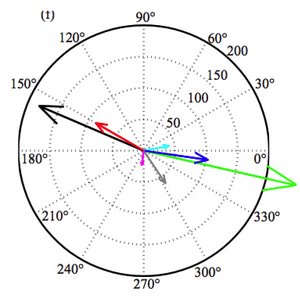
Our ears are able to detect sound intensities that vary over twelve orders of magnitude. This extraordinary ability of our sense of hearing relies on active amplification based on nonlinear dynamic oscillators in the ear. While the general principles of this cochlear amplifier have been characterized, the specific implementation is still unclear. Here we show that the interplay of hair bundle motility and electromotility provides a dynamic system that could be the basis of the cochlear amplifier.
D. O Maoileidigh and F. Jülicher
J. Acoust. Soc. Am., 128, 1175 (2010)
[PDF (592 kB)]
Intercellular Coupling Regulates the Period of the Segmentation Clock
The segmented organization of vertebrate animals is established by the collective dynamics of genetic cellular oscillators. We show that a simple model based on time delayed coupling of neighboring oscillators can account for the dynamic patterns observed in wild type zebrafish and in mutants. We show that the oscillation period is modified in mutants in which the time delay is altered. Furthermore, we observe the signatures of a dynamic instability in the correlation functions of oscillating patterns in mutants with reduced time delays. Such instabilities are predicted by theory and can be studied in numerical simulations.
L. Herrgen, S. Ares, L. G. Morelli, C. Schröter, F. Jülicher and A. C. Oates
Curr. Biol. 20, 1244 (2010)
[PDF (916 kB)]
Coupling a Sensory Hair-Cell Bundle to Cyber Clones Enhances Nonlinear Amplification
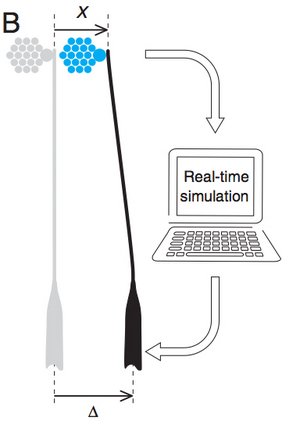
The ear posesses mechanosensitive hair cells which are able to detect minute vibtrations. These cells posess a hair bundle which is the sensory organelle. Hair bundles are active elements which can generate spontaneous movements and amplify periodic stimuli near a characteristic frequency. We have recently proposed that the amplification gain of hair bundles can be enhanced by elastically coupling of hair bundles in small groups. Here, we demonstrate this effect for a real hair bundle that is interacting with two "cyber bundles". These "cyber bundles" are virtual hair bundles which are simulated in real time on a computer.
J. Barral, K. Dierkes, B. Lindner, F. Jülicher and P. Martin
Proc. Natl. Acad. Sci. USA 107, 10765 (2010)
[PDF (395 kB)]
Bipedal Locomotion in Crawling Cells
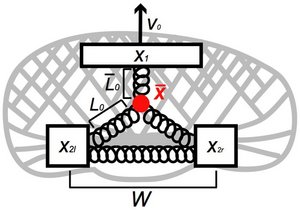
Keratocytes are motile cells that are able to glide on solid surfaces. Detailed inspection of their motility reveals that these cells exhibit an oscillatory modulation at the rear, reminiscent of bipedal locomotion. This locomotion can be understood as a slip-stick motion at the rear, induced by the gliding motion of the front part of the cell. Our theory can account for the observed changes in frequency of oscillations as a function of cell speed and adhesion strength.
E. L. Barnhart, G. M. Allen, F. Jülicher and J. A. Theriot
Biophys J. 98, 933 (2010)
[PDF (700 kB)]
High-Precision Tracking of Sperm Swimming Fine Structure Provides Strong Test of Resistive Force Theory
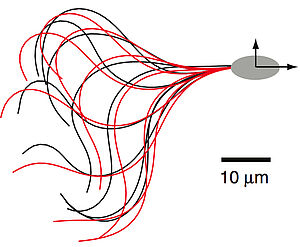
We quantify the shape of the ciliar beat of swimming sperm and analyze the fine-structure of the swimming trajectory. We show that the swimming path can be quantitatively understood using a resistive force theory. Our work reveals how the net curvature of the swimming trajectory emerges from asymmetries of the ciliar beat.
B. M. Friedrich, I. H. Riedel-Kruse, J. Howard and F. Jülicher
Journal of Experimental Biology 213, 1226 (2010)
[PDF (1,2 MB)]
Centrosome Size Sets Mitotic Spindle Length in Caenorhabditis Elegans Embryos
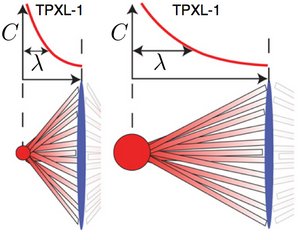
We show that the length of the mitotic spindle in C. elegans embryos is controled by the size of the centrosomes that are located at the spindle poles. The molecule TPXL-1 which is recruted on the centrosomes forms a graded concentration profile on the microtubules of the spindle. This observation suggests that length of the mitotic spindle might be regulated via a spatial gradient on the spindle.
G. Greenan, C. P. Brangwynne, S. Jaensch, J. Gharakhani, F. Jülicher and A. A. Hyman
Curr. Biol. 20, 353 (2010)
[PDF (544 kB)]